目录
硬脂酸如何对抗还原性压力
硬脂酸如何对抗还原性压力;香蕉奶昔研究 Redux
布拉德·马歇尔 / 2022 年 7 月 11 日
自从我发表《可松饮食》以来,人们对我称之为“香蕉奶昔研究”的科学文章产生了浓厚的兴趣^1^. 研究表明,在两天的低脂饮食后,在香蕉奶昔中摄入 24 克硬脂酸会导致线粒体融合和循环酰基肉碱的下降。

右图显示摄入硬脂酸增强香蕉奶昔后的线粒体融合。
我们对此非常感兴趣,因为喂食硬脂酸的大鼠已经腹肌明显。^2^

红色圆圈表示内脏脂肪。
香蕉奶昔研究的作者认为,线粒体融合是由于一种酶——TfR1——的硬脂酰化,该酶参与了影响线粒体融合的信号通路。我怀疑这是真的,但我也怀疑线粒体融合的发生是由于硬脂酸对还原性压力的影响——释放 NAD+。正如我喜欢说的,这些事情通常由多个强化反馈回路调节, 像乌龟一样慢。
在最近的帖子中,我认为肥胖是由还原性压力引起的。引起还原性压力的一种方法是将脂肪,尤其是不饱和脂肪,以比 NAD+ 再生更快的速度进入线粒体,从而导致 NADH 与 NAD+ 的比例很高(NADH 是还原形式,NAD+ 被氧化)。NAD+ 是脂肪燃烧的限制因素,当缺乏时,会积聚 NADH、乙酰辅酶 A 和循环酰基肉碱。这不好!
硬脂酸如何对抗还原性压力?
人们摄入的绝大多数膳食脂肪仅由四种脂肪酸组成:饱和硬脂酸和棕榈酸、单不饱和油酸和多不饱和亚油酸。其中三种——硬脂酸、油酸和亚油酸——是 碳18 脂肪,仅在不饱和度上有所不同。在这三种中,硬脂酸是迄今为止吸收和氧化最慢的。考虑摄入后前 9 小时内燃烧的三种脂肪的百分比^3^:

膳食硬脂酸几乎被氧化(在线粒体中燃烧)。事实上,硬脂酸在各个层面上都是最慢燃烧的。当吃脂肪时,会被肠道包装成乳糜微粒,然后以游离脂肪酸的形式释放出来,这些脂肪酸通过转运蛋白 CD36 被细胞吸收,最终通过一种叫做 CPT1a 的酶转移到线粒体中,其表达受到PPAR α 控制。
第 1 步,吸收: 硬脂酸导致餐后乳糜微粒形成率低。^4^ 富含硬脂酸的混合物是图表中带有 x 和 S 的线。图表中的其他字母是亚油酸、肉豆蔻酸、油酸、棕榈酸和反式脂肪含量高的混合物。

硬脂酸作为乳糜微粒的吸收和释放比其他脂肪要慢得多。
第 2 步,以游离脂肪酸 (FFA) 的形式从乳糜微粒中释放:乳糜微粒“停放在”组织中,其中的脂肪以 FFA 的形式释放到血液中。释放它们的酶称为脂蛋白脂肪酶。与其他脂肪相比,LPL 对硬脂酸的活性较低。^4^

第三步,细胞摄取:CD36酶参与细胞摄取游离脂肪酸。其表达受 PPAR γ 的控制,PPAR γ 由油酸和亚油酸激活,但不受硬脂酸激活。 硬脂酸导致低水平的 CD36。^5^

与不同脂肪酸孵育的巨噬细胞中的 CD36 表达。
第四步,进入线粒体:控制脂肪进入线粒体速度的酶称为CPT1,其表达受PPARα控制。PPAR α 受油酸(C18:1) 刺激,但不受硬脂酸 (C18:0) 刺激。^6^
 C18:0 = 硬脂酸;C18:1 = 油酸;C16:1 = 棕榈油酸(16 碳MUFA,不常见);C16:0 = 棕榈酸;C14:0 = 肉豆蔻(14 碳饱和,存在于椰子油中);C12:0 = Lauric(椰子油)
C18:0 = 硬脂酸;C18:1 = 油酸;C16:1 = 棕榈油酸(16 碳MUFA,不常见);C16:0 = 棕榈酸;C14:0 = 肉豆蔻(14 碳饱和,存在于椰子油中);C12:0 = Lauric(椰子油)
步骤 4a,进入线粒体:PPAR α 控制 CPT1 的数量,但酶本身也有不同的偏好,喜欢穿梭进入线粒体的脂肪。 CPT1明显不喜欢硬脂酸。^7^
 阅读此表,说明列标签中的脂肪进入线粒体的速度比括号中的脂肪快 X 倍。因此亚麻酸(AKA α亚麻酸,n-3)进入线粒体的速度比硬脂酸(ST)快 12 倍。油酸(油酸)进入线粒体的速度是硬脂酸的 4.6 倍。棕榈酸进入线粒体的速度比硬脂酸快 11 倍。
阅读此表,说明列标签中的脂肪进入线粒体的速度比括号中的脂肪快 X 倍。因此亚麻酸(AKA α亚麻酸,n-3)进入线粒体的速度比硬脂酸(ST)快 12 倍。油酸(油酸)进入线粒体的速度是硬脂酸的 4.6 倍。棕榈酸进入线粒体的速度比硬脂酸快 11 倍。
第 5 步,重新释放:未被组织吸收的 FFA 被肝脏吸收,重新转化为甘油三酯并以 VLDL 的形式释放,其行为与乳糜微粒非常相似。 硬脂酸很少掺入 VLDL。^8^

从字面上看,在脂肪氧化途径的每一步,硬脂酸都是慢动作。 硬脂酸正在阻止脂肪大量流入线粒体,让它们跟上。相比之下,油酸——硬脂酸被SCD1酶迅速转化为——是一匹赛马。冬眠的动物会增加SCD1的表达以帮助变得迟钝,这有什么奇怪的吗?^9[^或者说脂肪组织中scd1](https://fireinabottle.net/glossary/scd1/)的表达与脂肪量 之间存在直接的相关性?^10^

我最近写了一篇关于大剂量亚油酸(另一种赛马)如何在短期内导致脂肪氧化增加,这限制了燃烧葡萄糖的能力,就像人肥胖一样。 II 型糖尿病的代谢标志实际上是在碳水化合物和脂肪的混合膳食以及大餐后脂肪氧化和低葡萄糖氧化后无法降低 FFA(又名 NEFA:非酯化脂肪酸)。^11^
考虑以下事实:天然SCD1抑制剂香苹婆油^12^,阻止硬脂酸转化为油酸,改善自发性肥胖大鼠的血糖控制。给予小鼠降低脂肪酶活性的药物^13^因此,FFA 水平具有出色的血糖控制能力。
硬脂酸减缓脂肪流动的所有这些加起来是什么?2021 年的一篇论文显示了硬脂酸对餐后 FFA 水平的影响。^14^ 胰岛素的主要工作之一是抑制脂肪分解以降低餐后 FFA 水平。这优先燃烧葡萄糖以防止血糖峰值。如果吃的是混合饮食,想在餐后抑制 FFA。与棕榈酸相比,硬脂酸可明显降低餐后 FFA,尤其是在连续进餐后。

我已经提到,植物油会急剧降低葡萄糖氧化,就像糖尿病患者一样。 硬脂酸似乎特别擅长允许餐后葡萄糖氧化。 硬脂酸会让人表现得像个瘦子。
[[https://fire-in-a-bottle.myshopify.com/a/bundles/fight-reductive-stress!-4871|对抗减轻压力 - 节省 10%!]]
这些产物共同作用以对抗还原性压力。硬脂酸减缓脂肪流入线粒体。香苹婆油可防止硬脂酸转化为油酸。α-硫辛酸通过将 NADH 转化为 NAD+ 直接对抗还原性压力。
 *
*
消除还原性压力会导致线粒体融合。
但是,如果硬脂酸实际上减慢了脂肪氧化的速度,那么为什么硬脂酸会导致线粒体融合、循环酰基肉碱减少和“脂肪燃烧模式”呢?这是一个悖论吗?
想成为乌龟,还是想成为兔子?
这是我对香蕉奶昔研究的看法。由于大多数美国人储存的油酸和亚油酸含量很高,因此当他们进行两天的低脂纯素饮食时,参与者可能会经历减少的压力。低 NAD+ 水平与线粒体碎片有关。^15^当他们食用香蕉奶昔时,会导致脂肪加工减慢,从而使线粒体恢复 NAD+ 水平,从而导致融合。此外,一些硬脂酸被燃烧,饱和脂肪通过 ROS/GR/NNT 途径产生更多的 NAD+。融合的线粒体具有更大的体积和表面积,线粒体酶没有被乙酰化,能够更有效地处理脂肪,最终导致循环中的酰基肉碱减少。降低混合餐后脂肪进入线粒体的速度,可以恢复参与者的 NAD+ 水平,使其进入脂肪燃烧模式。

NAD+ 前体烟酰胺单核苷酸可防止细胞因缺血而受到还原性应激(即氧气的短暂消除)后线粒体断裂。
结论
有很多证据表明,硬脂酸对线粒体融合和对抗腹部脂肪的影响与如何进入线粒体的动力学有很大关系。硬脂酸是脂肪中的乌龟。踏实和稳重是赢得比赛的关键。
减少餐后游离脂肪酸可以在恢复血糖控制方面发挥巨大作用,因为脂肪氧化通过兰德尔循环阻止葡萄糖氧化。将硬脂酸视为“脂肪纤维”并没有错。
Senyilmaz-Tiebe D、Pfaff DH、Virtue S 等。膳食硬脂酸调节人类体内的线粒体。纳通。2018年8月7日在线发表。doi:10.1038/s41467-018-05614-6
沉 MC、赵 X、Siegal GP、Desmond R、Hardy RW。膳食硬脂酸导致无胸腺裸鼠内脏脂肪组织减少。韦斯 SFT,编辑。公共科学图书馆一。2014 年 9 月 15 日在线发布:e104083。doi: 10.1371/journal.pone.0104083
Jones PJH、Pencharz PB、Clandinin MT。膳食脂肪酸的全身氧化:对能量利用的影响。美国临床营养学杂志。1985 年 11 月 1 日在线发表:769-777。doi: 10.1093/ajcn/42.5.769
Tholstrup T, Sandström B, Bysted A, Hølmer G. 6 种膳食脂肪酸对健康年轻男性餐后血脂、血浆脂肪酸、脂蛋白脂肪酶和胆固醇酯转移活性的影响。美国临床营养学杂志。2001 年 2 月 1 日在线发布:198-208。doi: 10.1093/ajcn/73.2.198
Vallvé JC、Uliaque K、Girona J 等人。不饱和脂肪酸及其氧化产物刺激人巨噬细胞中的 CD36 基因表达。动脉粥样硬化。2002 年 9 月在线发表:45-56。doi: 10.1016/s0021-9150(02)00046-1
Popeijus HE、van Otterdijk SD、van der Krieken SE 等。脂肪酸链长度和饱和度影响 HepG2 细胞中 PPARα 转录激活和抑制。Mol Nutr Food Res。2014 年 10 月 27 日在线发布:2342-2349。doi: 10.1002/mnfr.201400314
Gavino VC、Cordeau S、Gavino G. 大鼠线粒体中酰基肉碱合成选择性的动力学分析。脂质。2003 年 4 月在线发表:485-490。doi: 10.1007/s11745-003-1088-7
派T,叶YY。在培养的大鼠肝细胞中,硬脂酸对极低密度脂蛋白脂质组成和粒径的改变不同于短链饱和脂肪酸。脂质。1997 年 2 月在线发表:143-149。doi: 10.1007/s11745-997-0018-z
茶山 Y、安藤 L、佐藤 Y 等。叙利亚仓鼠冬眠之前和同时发生的白色脂肪组织重塑的分子基础,这是一种储存食物的冬眠者。前生理学。2019年1月28日在线发表。doi:10.3389/fphys.2018.01973
Caron-Jobin M、Mauvoisin D、Michaud A 等。肥胖女性腹部脂肪组织的硬脂酸含量。营养品 糖尿病。2012 年 1 月在线发布:e23-e23。doi: 10.1038/nutd.2011.19
Normand-Lauzière F、Frisch F、Labbé SM 等。2 型糖尿病患者餐后非酯化脂肪酸外观和氧化的增加在糖尿病受试者的后代中尚未完全确定。维拉 A,编辑。公共科学图书馆一。2010 年 6 月 4 日在线发布:e10956。doi: 10.1371/journal.pone.0010956
Ortinau LC、Nickelson KJ、Stromsdorfer KL 等。SCD1的天然抑制剂 Sterculic Oil可改善肥胖 OLETF 大鼠的代谢状态。肥胖。2013 年 2 月在线发布:344-352。doi: 10.1002/oby.20040
Schweiger M、Romauch M、Schreiber R 等。脂肪甘油三酯脂肪酶的药理抑制作用可纠正高脂饮食诱导的小鼠胰岛素抵抗和肝脂肪变性。纳通。2017年3月22日在线发布。doi:10.1038/ncomms14859
van Rooijen MA,Plat J,Zock PL,Blom WAM,Mensink RP。连续两次高含量棕榈酸或硬脂酸混合膳食对健康体重和超重男性和绝经后女性餐后 8 小时血脂和血糖的影响:一项随机对照试验。欧洲食品杂志。2021 年 3 月 17 日在线发布:3659-3667。doi: 10.1007/s00394-021-02530-2
Klimova N、Fearnow A、Long A、Kristian T. NAD+ 前体通过 SIRT3 依赖机制调节缺血后线粒体断裂和活性氧物质的产生。实验神经病学。2020 年 3 月在线发布:113144。doi: 10.1016/j.expneurol.2019.113144
硬脂酸与线粒体健康
当前反对多不饱和脂肪酸的运动最近受到关注。然而,饱和脂肪的好处在所有戏剧中常常被忽视。这是一种饱和脂肪的故事。
2022年8月23日
语境
最近,关于食用多不饱和脂肪酸(PUFA)(也称为种子油或植物油)对健康的影响引起了广泛的讨论。许多人已经从饮食中剔除多不饱和脂肪酸,转而选择更多的饱和脂肪酸 (SFA) 以及更有益的omega-3多不饱和脂肪酸。
尽管如此,当代争论的焦点似乎集中在omega-6 PUFA 的危害上。虽然这是一个值得讨论的问题,但SFA的好处也值得关注。
本文的重点是硬脂酸(一种长链 SFA)及其对线粒体的影响,最终影响能量代谢和整体健康。
实质

细胞器
线粒体是祖先原核细胞器,被认为是独立的有机体,在数十亿年前被真核(哺乳动物)细胞吞噬。
如果学过大学生物学入门课程,线粒体可能被视为细胞的动力源,但值得更多的描述。
线粒体对生命至关重要——人需要线粒体来产生大部分能量。线粒体在程序性细胞死亡、分子合成、钙调节、神经递质代谢中发挥作用,在氧化还原系统中发挥不可或缺的作用。

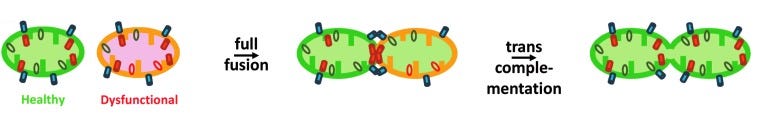
线粒体融合
两个互补的线粒体过程——融合和裂变——对于维持其形式和完整性至关重要。线粒体繁殖需要裂变,而融合则需要合作。
一、融合:
当两个或多个线粒体融合时,能够一起工作并更有效地产生能量。虽然碎片化的线粒体可能没有任何产生能量的问题,但两个融合的线粒体可以在整个隔室中共享彼此的资源(代谢物、酶等)。
线粒体融合还可以通过一种称为反式互补的过程来纠正功能失调的基因突变,其中一个缺乏功能基因的线粒体可以与另一个含有相同基因的健康副本的线粒体融合。有缺陷的 mt-DNA 副本可以被纠正或补偿,由此产生的融合线粒体可以恢复功能。
饱和脂肪酸
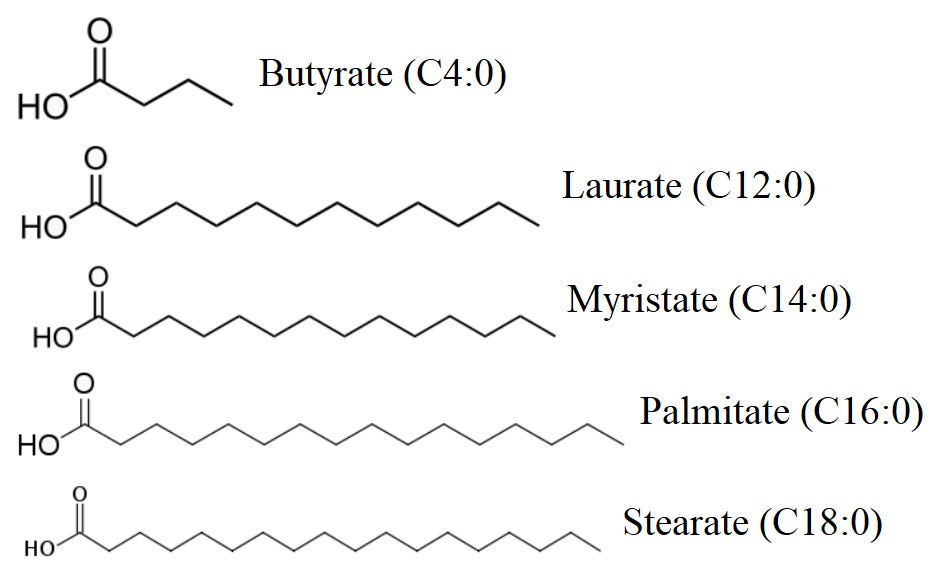
SFA 是含有甘油主链和仅碳-碳单键的脂肪酸,具有最大数量的氢原子。
在过去的一个世纪里,SFA 被广泛妖魔化为心血管和代谢疾病的驱动因素。然而,最近的研究已经消除了 SFA 长期以来所认为的后果。
看看我们之前的一篇文章,讨论了饱和脂肪的稳定性以及 PUFA(多不饱和脂肪酸)在慢性疾病中的作用:
硬脂酸,或 C18:0,是一种具有 18 个碳链的饱和脂肪酸 (SFA)。硬脂酸的重要膳食来源包括牛油、猪油、黄油、肉类、家禽、鱼类和奶制品。
一个有趣的实验
为了研究硬脂酸的影响,研究人员进行了一项研究,受试者(健康/糖尿病)采用两天低脂纯素饮食,以降低硬脂酸基线,然后喝高硬脂酸饮料。
为期 2 天的低脂纯素饮食(C18:0 含量极低)旨在确保受试者几乎没有循环 C18:0。随后,他们喝了一杯含有24克硬脂酸的香蕉奶昔饮料。
在喝奶昔前后,对受试者的血液进行线粒体断裂和融合测试。为了方便起见,选择观察中性粒细胞(一种白细胞)。
经过两天的低脂饮食后,发现中性粒细胞线粒体相当破碎。约50%的线粒体断裂,<10% 融合。这种效果在所有 21 名受试者中都是一致的。
饮用富含硬脂酸的混合物3-6 小时后诱导线粒体融合。碎片率下降至 25%,而融合率则上升至 27%。受试者的结果再次稳健。
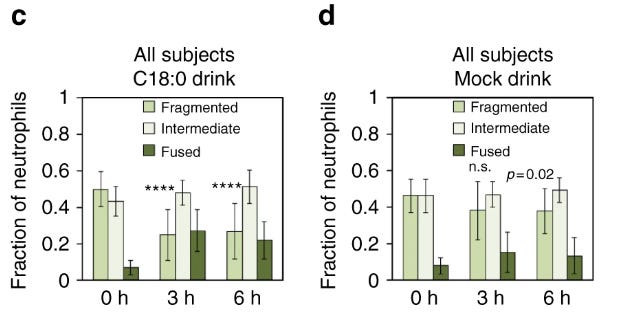
此外,为了确保饮料中只有 C18:0(而不是其他成分)负责线粒体融合,一名受试者服用了 C18:0 水乳液。线粒体如预期融合。
这些数据强烈表明 C18:0 快速引起人类中性粒细胞中的线粒体融合。据推测,其他人类细胞也可能通过类似的过程进行线粒体融合。
作者进一步测试了线粒体融合对另一种饱和脂肪——棕榈酸(C16:0) 的反应。在喝了等量 C16:0 的饮料后,受试者的细胞中没有表现出线粒体融合。其他研究也发现 C18:1(油酸)和 C20:0(花生酸)等脂肪也是类似的效果。
关于酰基肉碱的旁注
肉碱是一种参与能量代谢的分子,有助于将长链脂肪酸转运到线粒体中进行氧化,最终产生能量。酰基肉碱是乙酰辅酶A和肉碱的复合物,可将脂肪酸输入线粒体。
线粒体中发生的氧化类型是β-氧化。这个过程将脂肪酸分解成参与电子传输链的分子,这是有氧产生能量 (ATP) 的主要机制。
- YouTube")
研究人员测量了受试者摄入硬脂酸奶昔前后血液中酰基肉碱的水平。两天低脂饮食后,血清中酰基肉碱水平较高,但饮酒后则明显下降。
低脂饮食后血液中酰基肉碱的存在,可能与线粒体功能障碍导致的β-氧化受损有关。另外,低脂饮食可能意味着碳水化合物是这些人的主要能量来源,这意味着肉碱会积累并未被利用。
虽然饮酒后酰基肉碱水平下降,但循环中的C18:0 三酰基甘油酯却飙升。这种关系表明酰基肉碱水平的下降是对线粒体β-氧化的需求的响应。
再次测试了其他脂肪酸,例如 C16:0(棕榈酸酯)。然而,血清酰基肉碱水平没有受到影响,这表明硬脂酸具有特定的作用。
**为什么线粒体功能障碍很重要**
当线粒体过度破碎时(意味着融合存在缺陷),融合调节蛋白很可能会被禁用。干扰线粒体融合的过程会引起氧化应激、代谢问题和整体线粒体功能障碍。
线粒体功能障碍可以遗传,也可以通过生活方式诱发。这具有临床意义,因为胰岛素抵抗引起的慢性疾病的病理生理学涉及线粒体融合缺陷。值得注意的例子包括:
- 2 型糖尿病 (T2DM)
- 神经退行性疾病(阿茨海默症、帕金森氏症)
糖尿病
2型糖尿病(T2DM) 涉及线粒体功能障碍、氧化剂产生和ATP产生受损。对小鼠进行的一项研究表明,融合蛋白的缺陷(过度断裂)会损害葡萄糖稳态,导致胰岛素抵抗和肥胖。
此外,糖尿病患者的胰腺β细胞的融合/裂变调节不断受到损害。例如,高血糖会抑制融合,阻止线粒体 O2 消耗。
线粒体融合蛋白 2 蛋白表达降低与 T2DM 相关。在 T2DM 患者的白细胞中,观察到线粒体分裂增多,融合减少。
运动已被证明可以通过增加融合蛋白和减少裂变蛋白来提高胰岛素敏感。
许多研究表明 T2DM 存在线粒体分裂的倾向。
综上所述
基于以上所述,我不希望您相信线粒体裂变导致代谢疾病的论点。
我希望你明白的是……先有鸡还是先有蛋……这并不重要。重要的是了解是什么导致线粒体承受更大的压力……和低程度的支持。
从生理学角度来看,无论是什么原因导致糖尿病或慢性代谢疾病的后果……也与削弱线粒体功能的表型有关。
因此,读者有必要参加增强线粒体功能的活动。
优化线粒体功能的营养素对于维持身体的能量平衡很重要,应包含在饮食中。硬脂酸是这些营养素之一,可以从肉、鱼、家禽和黄油中获取。硬脂酸对脂肪酸氧化和线粒体融合的潜在益处不容忽视。
https://www.remnantmd.com/p/stearic-acid-and-mitochondrial-health
硬脂酸作为信号分子:SEA硬脂酰乙醇酰胺
Brad Marshall / 2023 年 6 月 4 日
https://www.youtube.com/watch?v=X-0h8XOQgII
在最新的视频中,我解释了乙醇酰胺系统的基础知识。您可能听说过 OEA,一种由油酸(橄榄油)制成的乙醇酰胺。OEA 可以降低热量摄入和炎症。
乙醇酰胺可以由任何脂肪制成。当由 欧6 花生四烯酸制成时,会变成 AEA ,并触发大麻素受体。AEA 与肥胖和暴饮暴食有关。
SEA(硬脂酰乙醇酰胺)由硬脂酸制成。在主要的乙醇酰胺中,SEA 研究最少,已被证明可以降低食欲、抑制肝脏中的 SCD-1,对减少炎症具有明显效果。还被证明可以恢复胰岛素抵抗大鼠的 Delta 6 去饱和酶 (D6D) 活性水平。正如我在上周的视频中指出的,去饱和酶可以预测代谢疾病的进展。
SEA 与其他乙醇酰胺略有不同 - 不会触发大麻素受体或 PPAR α。有一些证据表明其控制与 OEA 相反。我在视频中讨论了为什么可能希望获得乙醇酰胺的益处,而不触发 PPAR α 或大麻素受体。
我在《身体脂肪成分的历史》中指出,在过去的80年里,硬脂酸在体脂中急剧下降,而且是独一无二的。我在这段视频中提供的证据表明,这可能导致循环上升水平必然下降。
我的推测是,硬脂酸补剂的一些神奇作用是通过 SEA 的上调来实现的。
https://fireinabottle.net/stearic-acid-as-a-signalling-molecule-sea-stearoylethanolamide/
体脂成分的历史
布拉德·马歇尔 / 2023 年 5 月 21 日
https://www.youtube.com/watch?v=YaM0KJzaZqU
下面是最新视频的摘要:
现在生来与过去不同。过去 80 年来,美国人体脂成分发生了巨大变化。第一份可靠数据于 1943 年发布。

1943 年,美国人体脂大约为 36% 饱和脂肪、43% 单不饱和脂肪和 10% 多不饱和脂肪PUFA普发。去饱和酶指数 (DI18)——油酸 (18:1) 与硬脂酸 (18:0) 的比率——为 6.6。到 1998 年,这些比率变为 24/55/21 和 16.2。

早在1964年,英国一项研究就报道称,心脏病和肥胖症患者的去饱和酶指数会升高。当时英国人的去饱和酶指数比其他地方报道的要高:

体脂成分由多种因素决定:饮食和酶。脂肪生成(“脂肪制造”)酶从其他能源中产生饱和脂肪。然后,这种饱和脂肪可以通过称为 SCD-1 的脂肪生成酶转化为油酸。当脂肪生成酶表达非常高时,油酸水平就会升高。饮食中的不饱和脂肪会导致脂肪生成酶上调,从而导致饱和脂肪降低和单不饱和脂肪酸升高,这是一个正反馈循环。高度不饱和的体脂会导致脂肪生成酶的持续上调。这可以通过实验室中的小鼠来证明:

这就导致了膳食PUFA增加导致脂肪生成酶增加的情况。这些脂肪生成酶有利于 MUFA 的产生。这意味着,随着体脂中 PUFA 含量的增加,饱和脂肪含量下降,而 MUFA 水平基本保持不变。如果将 1998 年研究中的三种 18:1 (油酸的不同版本)加在一起,油酸实际上比 1943 年报道的要高一些。
80 年来,与 SFA 相比,MUFA 和 PUFA 的水平有所上升。
https://fireinabottle.net/the-history-of-bodyfat-composition/
