糖尿病、危险脂肪和保护糖
2013 年 3 月 8 日,星期五
糖尿病、危险脂肪和保护糖
自 1963 年发现所谓的“葡萄糖-脂肪酸循环”以来,越来越多的证据表明游离脂肪酸与糖尿病有关(Randle、Garland、Hales 和 Newsholme,1963)。简而言之,葡萄糖-脂肪酸循环描述了一种竞争,即葡萄糖的使用会因脂肪酸的存在而受损,反之亦然(Cook、King 和 Veech,1978 年)。
其他人之前曾暗示过这种脂肪酸诱导的阻断作用。其中,有一位名叫Apollinaire Bouchardat的法国药剂师,给他的糖尿病患者喂食甜甜的水果和面筋制成的面包,效果不错。其他人应用了 Bouchardat 的饮食处方并取得了同样的成功。最近,跟随这些先驱的英国医生威廉·巴德 (William Budd) 每天给他的糖尿病患者服用约 8 盎司的糖,目的只是为了减缓长期控制不佳的糖尿病所特有的恶病质。他的大多数患者不仅停止消瘦,而且在接受这种“糖精治疗”后,他们的尿液中的糖分也开始停止流失(休斯,1862 年)。
另一位著名的临床医师兼研究员 Harold Himsworth 也是第一个证明糖尿病患者组织中的胰岛素敏感性降低的人,几十年后,根据他的临床经验和对人群数据的回顾,他建议高摄入膳食脂肪。游离脂肪酸水平升高)导致糖尿病,而富含碳水化合物和低脂肪的饮食对其有保护作用(Himsworth, 1934a, 1934b, 1936)。 卡恩等人。此后,通过多变量分析,实际上再次证实了希姆斯沃斯收集的数据,表明吃糖多的男性比吃少糖的人更不容易患糖尿病——信不信由你(卡恩等等,1971)。到 1980 年代,糖几乎已经从推测中归于它的疾病中免除。 这些研究人员的经历与大多数当代临床医生和非专业人士所坚信的观念不一致,即糖尿病是一种血液中葡萄糖或胰岛素过多的疾病。如果是这种情况,限制碳水化合物或服用将血糖或胰岛素水平降低到正常水平的药物,应该可以 180 度改变糖尿病的病程。 三项相对较新的临床试验的结果虽然远非完美,但削弱了对这种方法的信心。简而言之,在试验中,老年糖尿病患者被分配接受标准护理或更强化的治疗方案,其中通过各种方式将血糖水平降低至正常或接近正常水平,以降低血糖水平。死于中风和心脏病。总之,在血糖水平降至正常水平最多的受试者中,所有原因导致的死亡人数更多(Gerstein 等人,2008 年;Patel 等人,2008 年;Duckworth 等人,2009 年)。 (人们立即想到了所谓的“胰岛素增敏剂”罗格列酮,它现在带有一个关于心血管风险增加的黑框警告。) 这就引出了一个问题,如果碳水化合物和糖不是糖尿病的原因,如果血糖和胰岛素水平升高也不是,博主和非常受欢迎的健康和饮食网站之间散布的想法,究竟是什么? ? 考虑到糖尿病的初期缺陷——即胰岛素抵抗——之前描述的葡萄糖-脂肪酸循环,并吸收 Bouchardat 等人的临床经验,我们将首先转向酶复合物丙酮酸脱氢酶( PDH) 在探索这个问题。 PDH 复合物是一种至关重要的酶,它通过细胞质中的糖酵解和线粒体中的克雷布斯循环连接葡萄糖代谢。我在别处讨论过,葡萄糖的主要抑制发生在这里,而不是葡萄糖通过质膜穿梭进入细胞,例如 (Randle & Smith, 1958a, 1958b)。 脂肪酸直接和间接地使 PDH 复合物失活,阻止葡萄糖代谢为二氧化碳,而碳水化合物则具有相反的作用。糖尿病患者血液中的游离脂肪酸水平通常升高,这说明了他们的许多过程脱轨,包括 (1) 从一种燃料来源(例如脂肪)转换到另一种燃料来源(例如,葡萄糖)和(2)与非糖尿病患者相比,心血管疾病风险增加2 至 4 倍。 PDH 复合物的抑制阻止了丙酮酸代谢为乙酰辅酶 A,这是碳水化合物、脂肪和蛋白质代谢的汇聚点(图 1)。 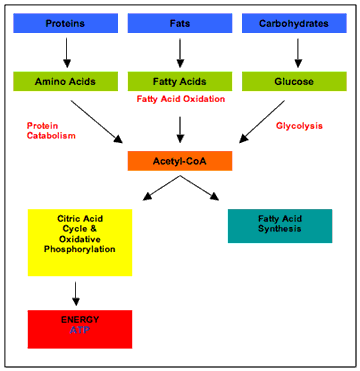
图 1代谢概述
这反过来又积压了 PDH 复合物上游的糖酵解酶。结果,游离葡萄糖分子在细胞中积累,并随着它们的浓度梯度移回血液中(Sonksen,2001)。 PDH 不仅受脂肪酸抑制,而且受其氧化产物抑制(Bowker-Kinley、Davis、Wu、Harris 和 Popov,1998)。相反,葡萄糖氧化提供丙酮酸,后者直接刺激 PDH 活性,并激活(并增加表达)乙酰辅酶 A 羧化酶,乙酰辅酶 A 羧化酶是一种通过生成丙二酰辅酶 A 阻止脂肪酸氧化的酶(Bouzakri 等,2008) (图2)。后一个过程被称为羧化反应,因此,葡萄糖氧化产生的二氧化碳比脂肪酸氧化产生的二氧化碳量更大,从而推动反应向前发展。 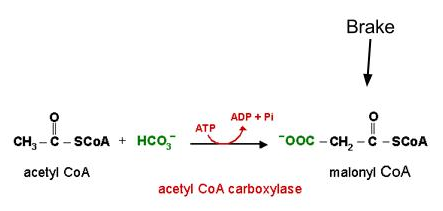 图 2由葡萄糖氧化产生的丙二酰辅酶 A 对脂肪酸氧化施加了“刹车”。 PDH 受到高度调控,既不改变也不转录较少。相反,PDH 更有可能被各种因素保持在非活性状态,即游离脂肪酸、酮体和乳酸 - 根据导致发现葡萄糖-脂肪酸循环 (McCromack, Edgell, & Denton, 1982)。
图 2由葡萄糖氧化产生的丙二酰辅酶 A 对脂肪酸氧化施加了“刹车”。 PDH 受到高度调控,既不改变也不转录较少。相反,PDH 更有可能被各种因素保持在非活性状态,即游离脂肪酸、酮体和乳酸 - 根据导致发现葡萄糖-脂肪酸循环 (McCromack, Edgell, & Denton, 1982)。
尽管脂肪酸和葡萄糖都会产生乙酰辅酶A,但相似之处仅止于此。
一方面,葡萄糖氧化降低了能量 (ATP) 产生的氧气成本,而 ATP 通过更具推测性的机制调节葡萄糖进入细胞的过程。脂肪酸的氧化比葡萄糖的氧化消耗更多的氧气,因此导致其中的氧张力较低。 氧张力的降低反过来刺激了分泌胶原蛋白的成纤维细胞的增殖,而这种新铺设的胶原蛋白通过延长氧气扩散到达细胞的距离来进一步降低氧张力。此后,成纤维细胞以典型的正反馈方式沉积更多的胶原蛋白等,这最终可能导致更大程度的动脉粥样硬化,因为与其他脂质相比,胆固醇需要最大量的氧气进行氧化。 将优先燃料使用从脂肪酸转移到葡萄糖(其中一些正在研发中)的药物已被证明对糖尿病患者具有高度保护作用,并且副作用是心血管疾病。这是因为 ATP 的产生具有更高的氧气经济性。由 ATP 快速消耗引起的代谢压力可以超越葡萄糖 - 脂肪酸循环,在这种情况下,葡萄糖将转化为乳酸而不是二氧化碳(Bergeron 等,1999;Young, Radda, & Leighton, 1996)。 另一方面,目前使用的口服“降血糖”药物会带来中风和心脏病发作导致的死亡副作用(1970 年代由大学糖尿病研究小组首次发现)——这些事件的任务是降低血糖水平一开始就进行了。(这些药物(不是胰岛素)可以预防心血管疾病的证据也很少。) 第二,脂肪酸向组织的过度输送,作为一种保护性反射,增加了其中解偶联蛋白的表达,导致 ATP 的产生减少(以及随后的组织功能受损)。例如,在胰腺中,解偶联蛋白的表达增加,通过消耗 ATP,保持电压门控钙通道关闭,这是分泌胰岛素所必需的(图 3)(Zhang 等人,2001 年) )。 
图 3解偶联蛋白 (UCP) 是将来自葡萄糖的电子作为热量耗散而不是用于生成 ATP 的手段。需要 ATP 来刺激胰腺 β 细胞分泌胰岛素(Zhang 等,2001)。 三,葡萄糖氧化,比脂肪酸氧化更多,导致更多的气体,二氧化碳的产生。结果,氧气从红细胞向组织的卸载变钝(根据玻尔效应),进一步降低了氧气水平(图 4)。(这反映在呼吸商数中。)通常,糖尿病患者排出的二氧化碳比非糖尿病患者少(Simonson、Tappy、Jequier、Felber 和 DeFronzo,1988)。 .gif)
.png) 图 4 ** 上图:血红蛋白的脱氧状态由细胞的酸度 (H+) 维持,即通过产生二氧化碳 (CO2),使血红蛋白上的胺 (NH2) 基团羧化(未显示),从而增强其酸度,等等它们与血红蛋白分子的其他带正电部分形成盐系的能力。结果,血红蛋白的氧分子被释放到组织中。 底部:盐键(红色波浪线)稳定血红蛋白(T 结构)的脱氧状态。盐键的丢失使血红蛋白能够以更大的亲和力(R 结构)结合氧分子。 第四,脂肪酸高氧化率所固有的一个后果是细胞中脂肪酸代谢物(如神经酰胺和二酰基甘油)的积累,最终通过汇聚于激活的途径损害所谓的“胰岛素受体”。蛋白激酶 C (Erion & Shulman, 2010)。 五,较低的呼吸商伴随着较低的细胞能量电荷,仅这一点就可以解释糖尿病中的许多脱轨过程。例如,当细胞中的能量电荷减少时,钠进入细胞(降低血液中的钠含量),细胞保留钾的能力减弱。这可以解释为什么糖尿病患者的醛固酮水平高于平均水平,肾上腺皮质的醛固酮分泌由钠浓度或钾浓度下降触发(图 5)(Hollenberg 等人,2004 年;Kraus 、Jäger、Meier、Fasshauer 和 Klein,2005 年;Maalouf、Cameron、Moe 和 Sakhaee,2010 年)。
图 4 ** 上图:血红蛋白的脱氧状态由细胞的酸度 (H+) 维持,即通过产生二氧化碳 (CO2),使血红蛋白上的胺 (NH2) 基团羧化(未显示),从而增强其酸度,等等它们与血红蛋白分子的其他带正电部分形成盐系的能力。结果,血红蛋白的氧分子被释放到组织中。 底部:盐键(红色波浪线)稳定血红蛋白(T 结构)的脱氧状态。盐键的丢失使血红蛋白能够以更大的亲和力(R 结构)结合氧分子。 第四,脂肪酸高氧化率所固有的一个后果是细胞中脂肪酸代谢物(如神经酰胺和二酰基甘油)的积累,最终通过汇聚于激活的途径损害所谓的“胰岛素受体”。蛋白激酶 C (Erion & Shulman, 2010)。 五,较低的呼吸商伴随着较低的细胞能量电荷,仅这一点就可以解释糖尿病中的许多脱轨过程。例如,当细胞中的能量电荷减少时,钠进入细胞(降低血液中的钠含量),细胞保留钾的能力减弱。这可以解释为什么糖尿病患者的醛固酮水平高于平均水平,肾上腺皮质的醛固酮分泌由钠浓度或钾浓度下降触发(图 5)(Hollenberg 等人,2004 年;Kraus 、Jäger、Meier、Fasshauer 和 Klein,2005 年;Maalouf、Cameron、Moe 和 Sakhaee,2010 年)。

图 5血液中钾 (K+) 和钠 (Na+) 水平对醛固酮 (Aldo) 分泌的调节。 醛固酮与许多不良后果有关,包括炎症、血管问题、胰岛素敏感性下降以及用疤痕组织替代健康组织。瞧,拮抗醛固酮(盐皮质激素)受体的药物已被证明可以减缓糖尿病并发症的进展(Bender、McGraw、Jaffe 和 Sowers,2013 年)。醛固酮的作用与胰岛素的作用不一致,因为胰岛素具有促代谢作用,而醛固酮具有抗代谢作用。 六、脂肪酸氧化增加了乳酸和质子积累的趋势,这两者与游离脂肪酸同时升高,并且在肥胖和糖尿病中(Chen、Varasteh 和 Reaven,1993 年;Reaven、Hollenbeck、Jeng、Wu, &陈,1988)。与游离脂肪酸一样,乳酸会干扰胰岛素信号,增强胰岛素抵抗状态(Choi 等,2002;Depré、Veitch 和 Hue,1993)。 第七,脂肪酸的过度氧化,尤其是在氧化应激的背景下,可以通过从丙酮中产生晚期糖基化终末 (AGE) 产物(即通过其转化为甲基乙二醛)进一步损害组织的功能由于肝脏中脂肪酸的高氧化率。(感谢 Chris Masterjohn 博士为我澄清了这一点。)事实证明,高血糖在这个过程中可能扮演次要的或允许的角色。 所以总而言之,我的论点是脂肪酸的过度动员和氧化——一种糖尿病代谢的特征——会损害胰岛素作用,主要是通过抑制 PDH 复合物(Koves 等,2005)。简而言之,当游离脂肪酸水平升高时,葡萄糖转化为二氧化碳的代谢会受损,而葡萄糖会重新转化为乳酸。这对心血管疾病具有重要意义——总而言之,糖尿病患者患心血管疾病的风险增加。 那么,以我的迂回方式,如果您患有糖尿病或有糖尿病倾向,这如何转化为您现在可以开始申请的建议?这不是我的专业领域,实际上,其他地方的其他人在这方面做得很好。 尽管如此,我暂时建议,没有特别的顺序,(1)补充维生素 B1(PDH 复合物的辅因子)和(2)镁(有助于将 ATP 保留在细胞中),(3)像吃水果一样吃糖优先于谷物和块茎中的淀粉,(3) 减少体内多余的脂肪,并用肌肉代替,作为游离脂肪酸的吸收器,(4) 减少饮食中的脂肪,并用碳水化合物和蛋白质,保持卡路里或多或少相同。(我发现,而且大多数节食者都知道,肌肉的流失早在脂肪储存耗尽之前就发生了,这就是为什么在减肥期间碳水化合物优于脂肪的原因,因为碳水化合物具有很强的合成代谢作用),(5) 选择少食多餐,以在一天中保持稳定的血糖水平,(6) 尽可能减压一个论点,等等。这比执行计划本身更强大。. . 至少对我来说是这样。
参考 1. Randle, PJ, Garland, PB, Hales, CN & Newsholme, EA 葡萄糖脂肪酸循环。它在胰岛素敏感性和糖尿病代谢紊乱中的作用。柳叶刀1, 785-9 (1963)。2. Cook, GA, King, MT & Veech, RL 离体大鼠肝细胞的生酮和丙二酰辅酶 A 含量。生物化学杂志253, 2529–31 (1978)。3. Hughes, J. 医学科学改进和发现的季刊总结。美国医学杂志44, 232–235 (1862)。4. Himsworth, HP 影响葡萄糖耐量和胰岛素活性的饮食因素。生理学杂志81, 29–48 (1934)。5. Himsworth,HP 糖尿病管理。英国医学杂志2, 137–41 (1936)。6. Himsworth, HP 高碳水化合物饮食和胰岛素效率。英国医学杂志2, 57–60 (1934)。7. 卡恩、哈*等人。与糖尿病发病率相关的因素:对 10,000 名男性进行两年观察的多变量分析。以色列缺血性心脏病研究。慢性病杂志23, 617–29 (1971)。8. Gerstein, HC等。*强化降糖对 2 型糖尿病的影响。新英格兰医学杂志358, 2545–59 (2008)。9. 帕特尔,A.*等。*2 型糖尿病患者的强化血糖控制和血管结局。新英格兰医学杂志358, 2560–72 (2008)。10. Duckworth, W.*等。*2 型糖尿病退伍军人的血糖控制和血管并发症。新英格兰医学杂志360, 129–39 (2009)。11. Randle, PJ & Smith, GH 肌肉葡萄糖摄取的调节。1.胰岛素、厌氧菌和细胞毒物对离体大鼠膈肌摄取葡萄糖和释放钾的影响。生化杂志70, 490–500 (1958)。12. Randle, PJ & Smith, GH 肌肉葡萄糖摄取的调节。2. 胰岛素、厌氧菌和细胞毒物对糖类对离体大鼠隔膜渗透的影响。生化杂志70, 501–8 (1958)。13. Sonksen,PH 胰岛素,生长激素和运动。内分泌学杂志170, 13-25 (2001)。14. Bowker-Kinley, MM, Davis, WI, Wu, P., Harris, RA & Popov, KM 哺乳动物丙酮酸脱氢酶复合物组织特异性调节存在的证据。生化杂志329 ( Pt 1, 191–6 (1998))。15. Bouzakri, K.*等。*丙二酰辅酶 A 脱羧酶调节人类骨骼肌中的脂质和葡萄糖代谢。糖尿病57, 1508–16 (2008)。16. McCormack, JG, Edgell, NJ & Denton, RM 关于 Ca2+ 和丙酮酸在调节大鼠心脏丙酮酸脱氢酶活性中的相互作用的研究。饥饿和糖尿病的影响。生化杂志202, 419–27 (1982)。17. Bergeron, R.等。AMPK 激活对清醒大鼠肌肉葡萄糖代谢的影响。美国生理学杂志276, E938-44 (1999)。18. Young, ME, Radda, GK & Leighton, B. AICAR 激活大鼠骨骼肌中的糖原磷酸化酶和糖原分解——一种 AMP 活化蛋白激酶的激活剂。FEBS 信件382, 43–7 (1996)。19. 陈、CB等。β 细胞中解偶联蛋白 2 水平升高与葡萄糖刺激的胰岛素分泌受损有关:作用机制。糖尿病50, 1302–10 (2001)。20. 张,CY等。解偶联蛋白 2 对胰岛素分泌产生负面调节,是肥胖、β 细胞功能障碍和 2 型糖尿病之间的主要联系。细胞105, 745–55 (2001)。21. Simonson, DC, Tappy, L., Jequier, E., Felber, JP 和 DeFronzo, RA 胰岛素抵抗状态下果糖对碳水化合物诱导产热的标准化。美国生理学杂志254, E201-7 (1988)。22. Erion, DM & Shulman, GI 甘油二酯介导的胰岛素抵抗。自然医学16, 400–2 (2010)。23.霍伦伯格,NK等。糖尿病患者的血浆醛固酮浓度。肾脏国际65, 1435-9 (2004)。24. Kraus, D., Jäger, J., Meier, B., Fasshauer, M. & Klein, J. Aldosterone 抑制解偶联蛋白-1,诱导胰岛素抵抗,并刺激脂肪细胞中的促炎脂肪因子。激素和代谢研究 = Hormon- und Stoffwechselforschung = Hormones et métabolisme 37, 455–9 (2005)。25. Maalouf, NM, Cameron, MA, Moe, OW & Sakhaee, K. 2 型糖尿病低尿 pH 的代谢基础。美国肾脏病学会临床杂志:CJASN 5, 1277–81 (2010)。26. Bender, SB, McGraw, AP, Jaffe, IZ & Sowers, JR 盐皮质激素受体介导的血管胰岛素抵抗:糖尿病相关血管疾病的早期促成因素?糖尿病62, 313–9 (2013)。27. Chen, YD, Varasteh, BB & Reaven, GM 肥胖和 2 型糖尿病中的血浆乳酸浓度。糖尿病与代谢19, 348–54 (1993)。28. Reaven, GM, Hollenbeck, C., Jeng, CY, Wu, MS & Chen, YD NIDDM 患者 24 小时血浆葡萄糖、游离脂肪酸、乳酸和胰岛素的测量。糖尿病37, 1020–4 (1988)。29. Depré, C., Veitch, K. & Hue, L. 果糖 2,6-二磷酸在控制糖酵解中的作用。乳酸在离体工作大鼠心脏中刺激糖原合成。心脏病学报48, 147–64 (1993)。30. Choi, CS等。乳酸通过抑制糖酵解和削弱胰岛素信号传导,在骨骼肌中诱导胰岛素抵抗。美国生理学杂志。内分泌和代谢283,E233-40(2002 年)。31. Koves, TR等。过氧化物酶体增殖物激活受体-γ 共激活剂 1alpha 介导的骨骼肌细胞代谢重塑模拟运动训练并逆转脂质诱导的线粒体效率低下。生物化学杂志280, 33588-98 (2005)。 http://www.andrewkimblog.us/2013/03/diabetes-dangerous-fat-and-protective.html